
Skeletal Muscle Physiology
Overview
An Introduction to Muscle Types
The Neuromuscular Junction
Diffusion Affects Time of Signaling
Sarcomere Anatomy
How Ca2+, ATP, and Filaments Produce Muscle Contraction
Muscular Energetics: Contraction Depends on ATP Equilibrium
Muscle Types
An Introduction to Muscle Types

Figure 1. [3] There are several ways to classify muscles based on neural control, anatomy, or the presence of tissue striations. Skeletal muscle also falls under the category of voluntary muscle, whereas cardiac and visceral muscle fall under the category of involuntary muscles, or muscles that are not electively controlled. Skeletal and cardiac muscle can further be classified as striated and they both possess multiple nuclei, whereas visceral muscle falls under the category of smooth muscle because they possess singular nuclei.
The Neuromuscular Junction

Figure 2. [3]
A neuromuscular junction is shown in the diagram above. An action potential travels down the presynaptic nerve, causing voltage gated calcium channels (also known as voltage-dependent calcium channels or VDCC), to open. Because intracellular calcium concentrations are much lower than extracellular calcium levels, calcium ions to flow inward down their concentration gradient.
Calcium binds to synaptotagmin, a protein that binds acetylcholine (ACh)-filled vesicles to the presynaptic membrane. ACh is released via exocytosis and diffuses across the synaptic cleft and binds to acetylcholine receptors (AChR) on the postsynaptic muscle membrane. The binding of ACh to AChR, a ligand-gated ion channel (as opposed to a voltage-gated channel), enables sodium and potassium ions to flow inward, thereby depolarizing the muscle cell membrane. Once depolarization reaches threshold, an action potential is carried down the muscle cell, and propagates down the transverse tubule. Ion exchange pumps and SERCA proteins mediate the removal of excess calcium to turn off the signal. Acetylcholineesterase degrades acetylcholine into acetate and choline.
Figure 3. [3]
The diffusion of the signaling molecule acetylcholine across the neuromuscular synaptic cleft can be described by the equation above (left). The time for diffusion can be estimated when assuming one-dimensional diffusion where:
lambda = length of synaptic cleft (i.e., distance in µm)
D= Diffusion coefficient {µm2/s}
t= time of diffusion {s}
Though diffusion is not typically an effective means of transport, it is sufficient over small distances such as a synaptic cleft.
Flux in three dimensions represents a estimate of diffusion. Fick's Law of diffusion in three dimensions is shown on the right, where x, y, and z are coordinate directions, i, j, and k are directional unit vectors, J is flux {flow/unit area}, C is concentration {mol/L}, and D is the diffusion coefficient {area/time}.

Diffusion Affects Time of Signaling


Figure 4. [3]
In order to understand muscle contraction, we need to look at the individual myofibril, which is the individual unit within a muscle fiber; a myofibril is composed of bundles of myofilaments and it is this unit at which muscle contraction occurs. A myofibril is made up of thin filaments (composed primarily of actin) and thick filaments (composed of myosin), which vary their degree of overlap in order to produce muscle contraction. Within a sarcomere, the repeating unit within a myofilament (shown above), the portion consisting only of actin filaments is designated as the I-band. Within the I band there is a line called the Z-disks which are the end-to-end connection between actin filaments; there is a z-disk on each end of the sarcomere. There are two sets of myosin filaments on either side of the sarcomere midline (M-line). The entire region starting from one end of the myosin, past the midline, and to the opposite region of myosin is called the A-band. The H-zone is the region around the M-line where myosin filaments do not overlap.
Sarcomere Anatomy
How Ca2+, ATP, and Filaments Produce Muscle Contraction


Figure 5. [4], [3]. The overlap of actin and myosin filaments produces muscle contraction. As seen in the figure above, thin filaments are composed of G-actin, nebulin, troponin and tropomyosin. Tropomyosin blocks the myosin-binding sites on actin. Troponin is a complex of three proteins attached to the end of each tropomyosin molecule: troponin T, troponin C and troponin I. Troponin T binds to tropomyosin, troponin C binds calcium, and troponin I inhibits, or blocks the interaction between the myosin head group and its binding site on actin. The process of a myosin head group binding to actin is called cross bridge cycling. This process requires ATP. ADP and Pi bind to myosin, causing myosin to bind to actin. When ADP and Pi is released, the myosin head group undergoes a conformation change which rotates the head group 45°, known as the power stroke. ATP binds to myosin, decreasing its affinity for actin. ATP hydrolysis occurs and the myosin head group returns to its uncocked position, and troponin and tropomyosin re-cover the binding site on actin.
Muscular Energetics: Contraction Depends on ATP Equilibrium


Figure 6. [3] Muscular contraction during exercise is dependent on the continuous generation of ATP because of its role in altering the conformation of the myosin head group during cross bridge cycling. There are three major ways to regenerate ATP: creatine phosphokinase and myokinase regeneration, glycolysis, and oxidative phosphorylation. Creatine phosphokinase and myokinase have the fastest rate of ATP regeneration, but at low capacity. Glycolysis is the next fastest mode of ATP regeneration from pyruvate, but it does so at low capacity. Finally, oxidative phosphorylation produces the highest quantity of ATP per cycle but does so at the slowest rate.

Figure 7. 6 As mentioned earlier, muscle contraction is ATP-dependent. As a result, any process affecting ATP levels also affects the Na+/K+ ATPase pump, and through the subsequent altered sodium concentrations, the Na+/Ca2+ exchanger, and likewise the Ca2+pump. Altered ion concentrations and osmolarity affect fluid flow; therefore ion balance is important for maintaining cell size.
Muscle Types

Figure 8. [3] Type I muscle fibers are slow-twitch, oxidative (and consequently red), and fatigue-resistant fibers. On the other hand, type II fibers are fast twitch; they can either be oxidative and resistant or glycolytic and fatigable. Type 1 and IIa are utilized for aerobic endurance and some speed activities, whereas type IIb fibers are utilized for speed and power, such as in anaerobic activity (and are consequently white in comparison).

Figure 9. [3] As the figures above indicate, short duration activities require little ATP while long duration activities require more ATP. However, high intensity activities utilize ATP at a higher rate than low intensity activities. As mentioned earlier, type 1 and IIa fibers are used for endurance workouts, which pertains mostly to aerobic exercise such as long distance workouts. These particular muscle fibers produce slower velocity and therefore require a lower rate of ATP production. As a result, its main source of ATP comes from oxidative phosphorylation and glycolysis. Type IIb fibers, on the other hand, are utilized for speed and power, which pertains to anaerobic exercise, where oxygen does not have time to diffuse into the muscle. One example is sprinting. This activity and corresponding muscle type requires a high rate of ATP production and therefore its sources of ATP comes from creatine phosphokinase and glycolysis.
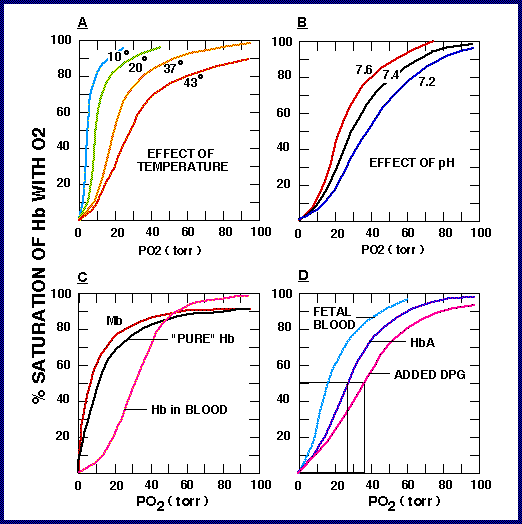
Figure 10. [5] Exercise requires a constant supply of O2 carried to the tissues by the blood. The oxygen saturation curves shown above describe the affinity of oxygen for hemoglobin when temperature, pH, DPG (also known as 2,3-Bisphosphoglycerate) are altered. Decreased pH, increased temperature, and increased levels of DPG all cause a shift to the right, decreasing oxygen-hemoglobin affinity. whereas increased pH, decreased temperature, decreased levels of DPG, and the presence of fetal hemoglobin all produce a shift to the left, increasing oxygen-hemoglobin affinity. Although not shown in the figure above, carbon dioxide levels result in a rightward shift, and carbon monoxide levels result in a leftward shift.




Figure 11. [2], [3]. The definitions of power, energy, and work can be combined into a single equation equating power to the product of force and velocity. The inverse relationship between force and velocity indicate that while heavier loads can create more force, it does so at the expense of a slower velocity.

Figure 12. [3] The diagram above is creating from experimental results from the relationship between power, force, and velocity as described in Figure 11. This curve indicates that fast-twitch muscles are more powerful due to their faster contractions. Slow twitch muscles have lower velocity, and therefore reduced power as compared to fast twitch muscles.